4.1
Bakterie
4.1.1
Morfologie
Souhrn
Bakterie jsou jednobuněčné prokaryotní mikroorganismy, které se morfologicky rozlišují na základní tvary:
- koky – diplokoky, tetrakoky, streptokoky, stafylokoky, sarciny
- tyčinky – rovné, zakřivené (vibria), kyjovité (korynebakteria), naštěpené (mykobakteria), pravidelné, nepravidelné
- vlákna – spirily, spirochety, vlákna se sporami (streptomycety)
Některé rody se vyznačují rozmanitostí tvaru buněk (tzv. pleomorfismus).
Velikost bakterií se měří v µm, šířka tyčinkovitých se pohybuje v rozmezí od 0,5 do 1,5 µm, délka je v rozmezí od 1 do 3µm, u téhož druhu je délka závislá na fyziologickém stavu buněk. Buňky, které se intenzivně rozmnožují, jsou mnohem kratší než buňky klidové.
Bakterie mohou být v různém uspořádání.
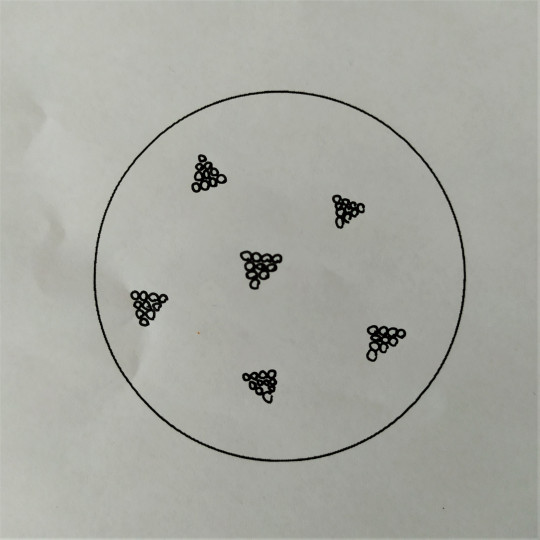
Koky mohou být ve dvojicích jako diplokoky, v řetízcích jako streptokoky, seskupené v hroznech jako stafylokoky, když jsou uspořádané po čtyřech tak jim říkáme tetrakoky. Tvoří-li útvary z osmi koků tvaru balíčků, nazýváme je sarciny. Všechno toto uspořádání závisí na tom, v jakých rovinách se děje dělení mikroorganismů.
+
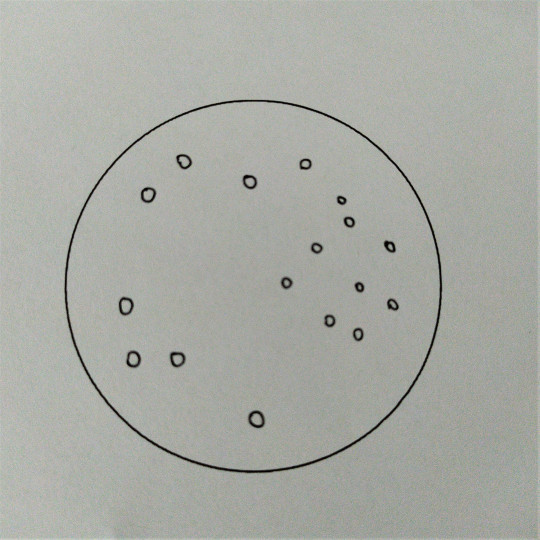
Obr. 11. Koky
+
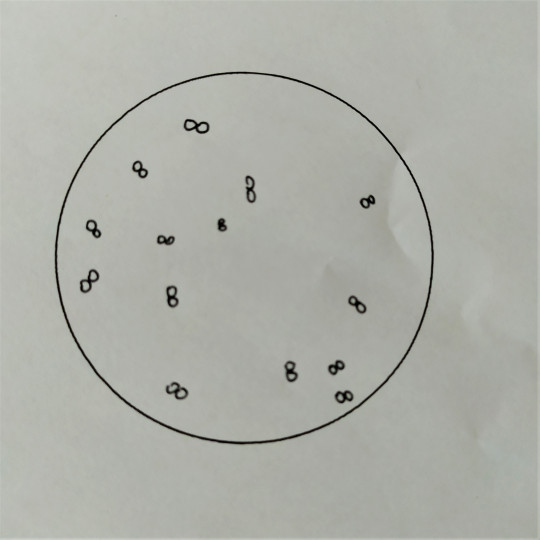
Obr. 12. Diplokoky
+
Obr. 13. Streptokoky
+
Obr. 14. Stafylokoky
+
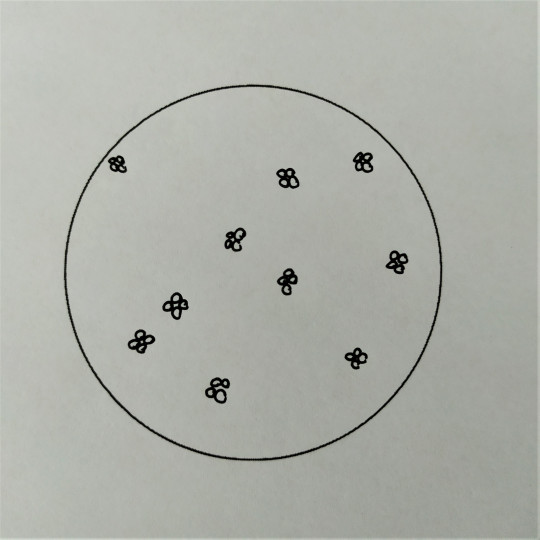
Obr. 15. Tetrakoky
+
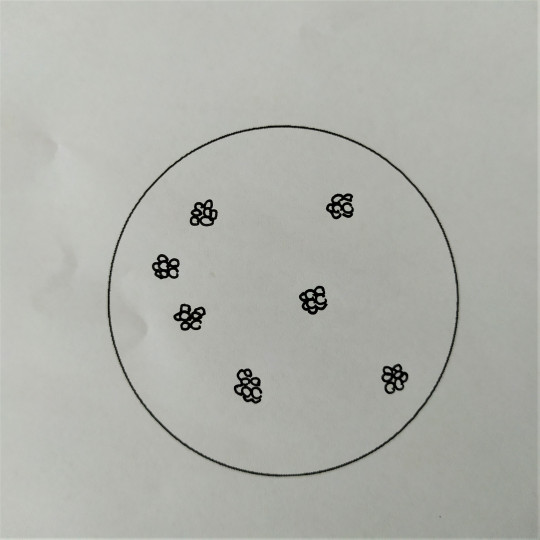
Obr. 16. Sarciny
+
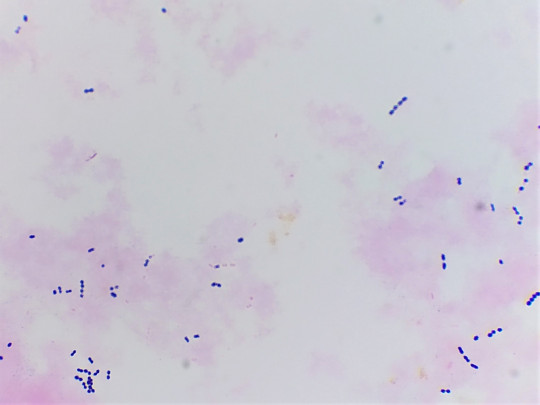
Obr. 17. Mikroskopický preparát – koky, diplokoky a krátké řetízky
+
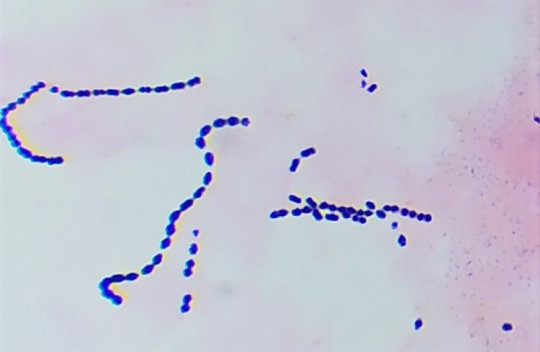
Obr. 18. Mikroskopický preparát – dlouhé řetízky
+
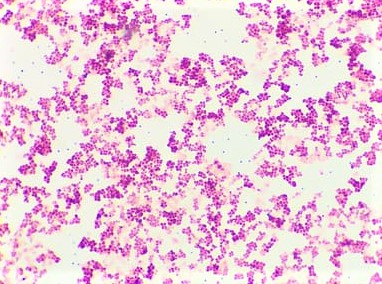
Obr. 19. Mikroskopický preparát - sarciny
Tyčinky mohou mít ještě více variací než koky. Rozlišujeme tyčinky krátké až kokovité, dlouhé až vláknité, štíhlé a silné s konci buď zaoblenými, rovně seříznutými, nebo naopak zašpičatělými až vzniká tvar vřetenovitý. Tyčinky mohou být stejnoměrně silné po celé délce, nebo nepravidelné, kyjovitě ztlustělé a různě zaškrcené. Sem nejčastěji řadíme bakterie a bacily. Rozdíl mezi bakteriemi a bacily je v tom, že bakterie netvoří spory, kdežto bacily ano. Spory jsou útvary, které bacily chrání před nepříznivými vnějšími vlivy. Dále mohou být tyčinky spirálovitého tvaru, tedy různě zahnuté. Říkáme jim vibria nebo spirily.
+
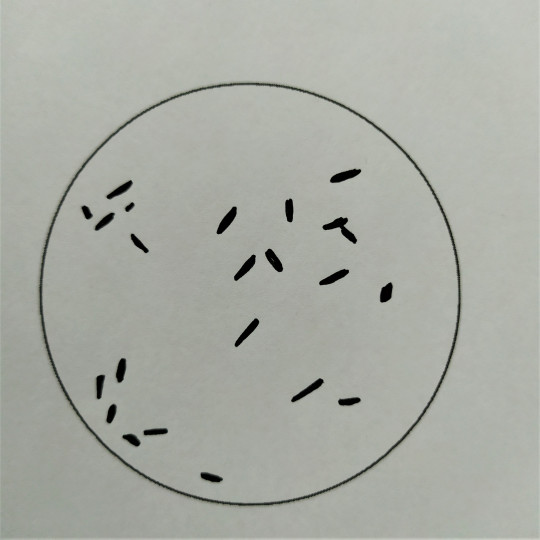
Obr. 20. Krátké tyčinky
+
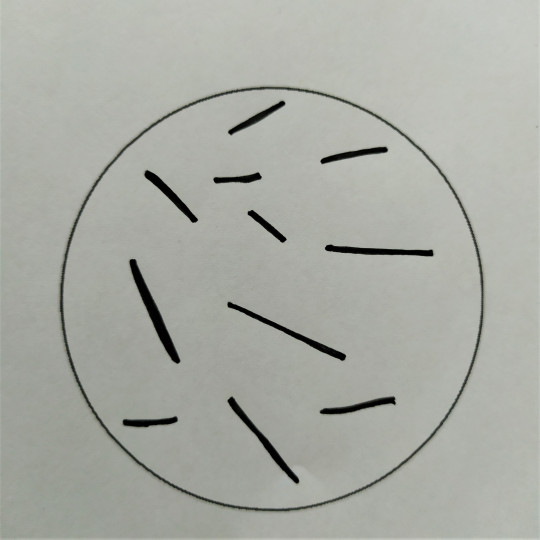
Obr. 21. Dlouhé tyčinky
+

Obr. 22. Tyčinky – vibria
+
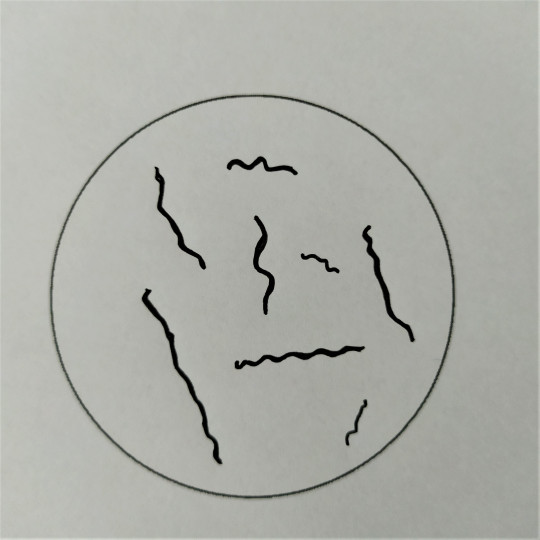
Obr. 23. Tyčinky – spirily
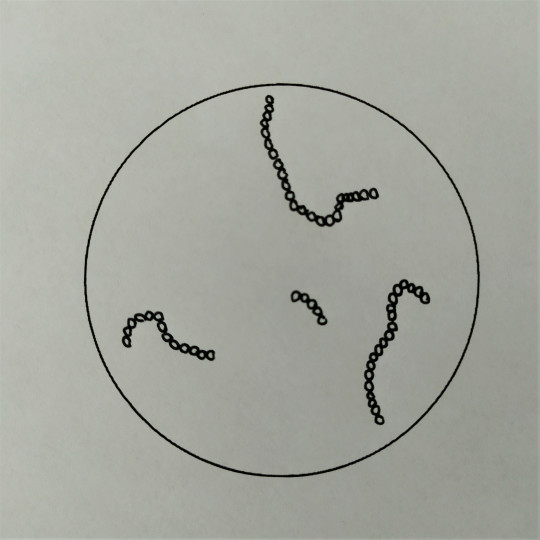
Vláknitý tvar tvoří přechod od bakterií k plísním. Vyskytuje se u aktinomycet. Vlákno může být hladké a pravidelné, nebo zrnité a nepravidelné. Může být větvené nebo nerozvětvené. Také vzájemné uspořádání vláken může být různé.
+
Obr. 24. Vláknité tyčinky
U téhož druhu se mohou tvarové vlastnosti bakterií měnit. Koky mohou měnit vzájemné seskupení. Tyčinkovité bakterie mohou měnit tvar jednotlivých tyčinek, například se může měnit jejich délka, šířka nebo zakončení. U různě zahnutých bakterií může dojít ke změně zakřivení. Vlivem nepříznivých vnějších podmínek mohou vznikat ve starých kulturách neobvyklé a nepravidelné tvary. Z těchto důvodů nemůžeme posuzovat a určovat bakterie výhradně podle velikosti a tvaru.
Jelikož jsou bakterie velmi malých rozměrů, musíme se někdy spokojit se studováním velkých souborů buněk. Takové soubory nebo populace buněk pěstujeme na laboratorních živných půdách. Živné půdy mohou být tekuté nebo tuhé. Celé tyto populace buněk nazýváme bakteriální kultury. Z čistých bakteriálních kultur po nárůstu na živné půdě zjišťujeme různé vlastnosti buněk. U směsných bakteriálních kultur, které jsou složeny z různých druhů mikroorganismů, posuzujeme jejich průmyslové uplatnění. Například kultury mlékárenské, pivovarnické, vinařské atd. Z každé buňky bakteriální kultury vzniká na živné agarové půdě jedna kolonie. Každá kolonie je tvořena souborem buněk, který pochází z jedné buňky a je tedy buněčným klonem. Daný druh nebo rod má za stálých podmínek kultivace stejný a charakteristický vzhled. U narostlých kolonií pozorujeme tvar, profil, okraje a konzistenci.
4.1.2
Cytologie
4.1.2.1
Buněčná stěna
Až na několik málo výjimek (mykoplazmata - buňky bez buněčné stěny) je na povrchu bakterie pevná a elastická buněčná (bakteriální) stěna. Buněčná stěna je propustná pro soli, nízkomolekulární a některé vysokomolekulární sloučeniny. Dává buňce tvar a chrání ji před mechanickými vlivy a před účinky osmotického tlaku vnějšího prostředí. V buněčné stěně jsou poměrně velké póry, kterými může volnou difúzí procházet většina chemických sloučenin. Vysokomolekulární látky, jako jsou polysacharidy nebo bílkoviny, nemohou póry buněčné stěny procházet. Hlavní stavební látkou buněčné stěny bakterií je peptidoglykan a murein.
Peptidoglykan obsahuje polysacharidová vlákna paralelně položená do řetězců, které jsou spojené tetrapeptidy nebo pentapeptidy. Peptidoglykan je tvořen N-acetylmuramovou kyselinou. Tloušťka peptidoglykanové vrstvy se u různých rodů bakterií liší. Působením enzymu lysozymu se peptidoglykany rozkládají a vzniká protoplast. Podle vrstvy peptidoglykanů v buněčné stěně se dělí bakterie na grampozitivní a gramnegativní.
Grampozitivní bakterie
Buněčná stěna grampozitivních bakterií se skládá z velmi silné peptidoglykanové vrstvy, která je vyplněná teikoovou kyselinou. Teikoové kyseliny mají lineární řetězce spojené s cytoplazmatickou membránou. Dále je na peptidoglykanovou vrstvu grampozitivních bakterií navázána řada polysacharidů. Ty jsou většinou složené z glukózy, galaktózy, manózy a řady dalších monosacharidů. Různé imunochemické reakce jednotlivých skupin bakterií způsobují právě polysacharidy.
Zajímavost
U grampozitivních bakterií v buněčné stěně nejsou žádné lipidy. Jen u mykobakterií a korynebakterií, jsou lipidy a vosky vázané esterovými vazbami na peptidoglykan.
Gramnegativní bakterie
Buněčná stěna gramnegativních bakterií je naopak tenká, ale složitější. Je složená z tenké vrstvy peptidoglykanů a je bez teikoové kyseliny. Nad touto tenkou vrstvou je vnější membrána podobná jiným biologickým membránám. Obsahuje fosfolipidy, bílkoviny a lipopolysacharidy. Mezi vnější membránou a peptidoglykanovou vrstvou je periplazmatický prostor. Ve fosfolipidové vrstvě jsou hydrofilní póry, tvořené bílkovinami zvané poriny. Tyto póry zasahují přes periplazmatický prostor až k peptidoglykanu, se kterým je pevně vázán.
Zajímavost
Ve stěně gramnegativních bakterií jsou i lipidy, které způsobují zvýšenou odolnost k aniontovým povrchově aktivním látkám, což mohou být mýdla nebo žlučové kyseliny. Díky této odolnosti se vyskytují gramnegativní bakterie ve střevním traktu savců. Stěnové lipopolysacharidy patogenů fungují jako endotoxiny, protože v těle živočichů vyvolávají charakteristické příznaky onemocnění.
Při přípravě protoplastu u gramnegativních bakterií působením lysozymu nedojde k úplnému odstranění všech složek buněčné stěny. Vzniklý útvar se proto označuje jako sféroplast a ten se mnohem snadněji regeneruje v normální buňku.
4.1.2.2
Cytoplazmatická membrána
Cytoplazmatická membrána se nachází pod buněčnou stěnou. Je jemná a elastická. Má malé póry. Obsahuje fosfolipidy a proteiny. Fosfolipidy mají dvojitou vrstvu, na okraji se nachází hydrofilní část a uprostřed je část hydrofóbní. Tato membrána vytváří vchlípeniny, které vybíhají do cytoplazmy. Zvláštním typem jsou mesozómy, které se vyskytují v oblasti, kde se při rozmnožování tvoří přepážka.
Cytoplazmatická membrána je polopropustná (semipermeabilní) a vytváří osmotické rozhraní buňky s vnějším prostředím. Polopropustnost zajišťuje, že póry volně prochází nedisociované molekuly vody, slabé kyseliny nebo alkoholy. Složkou lipidovou do buňky pronikají zase látky rozpustné v tucích. Pomocí zvláštních transportních mechanismů se do buňky dostávají všechny ostatní látky.
Poznámka
U fototrofních bakterií je v cytoplazmatické membráně obsažen také bakteriochlorofyl.
4.1.2.3
Bakteriální pouzdro
Mnoho bakterií má kromě buněčné stěny ještě bakteriální pouzdro. Bakteriální pouzdro se nachází vně buněčné stěny a je to vlastně slizovitý obal polysacharidové povahy (dextrany) nebo je z polypeptidů. Chrání buňku před vysycháním i před některými dalšími nepříznivými vlivy prostředí (zvýšená teplota, jedy, fagocytóza). Může tvořit sliz, což je dáno geneticky, ale také vlivem okolních podmínek (prostředí bohaté na cukry a chudé na dusíkaté živiny). Tvorba ohraničeného slizu se označuje jako kapsule nebo se difúzně sliz zřeďuje do prostředí, pak se označuje jako volný sliz. Pouzdra bakterií mají velmi úzký vztah k jejich životaschopnosti. Pod mikroskopem jsou pouzdra v neobarveném preparátu patrná jako jasná zóna kolem bakterie. V obarveném preparátu pouzdro barvu nepřijímá a zůstává neobarveno. Pouzdra se skládají z polysacharidů, polypeptidů, kyseliny hyaluronové a glutamové.
4.1.2.4
Cytoplazma
Cytoplazma tvoří vnitřní obsah buněk. Skládá se z buněčné šťávy, tedy koloidního roztoku globulárních bílkovin, v němž jsou rozpuštěny ribonukleové kyseliny, aminokyseliny, nukleotidy, soli organických kyselin, produkty látkové výměny a vitaminy. Cytoplazma vytváří vhodné prostředí pro funkci ribozomů.
Ribozomy jsou buněčné organely, v nichž probíhá syntéza bílkovin. Tvoří až 40 % sušiny cytoplazmy. Největší složkou cytoplazmy je voda, která představuje 65 až 90% obsahu buněk. Dále se u některých bakterií vyskytují barviva. Nejčastěji jde o karotenoidní (žlutá až červená) a melanoidní (černá), která nejsou uvolňována do prostředí. Řada dalších barviv je však vylučována do prostředí, jako např. fenazinová barviva.
4.1.2.5
Deoxyribonukleová kyselina
Jaderným materiálem u bakterií (prokaryotních) je deoxyribonukleová kyselina (DNA), která tvoří v buňce jedinou molekulu, představující chromozom, a je umístěna přímo v cytoplazmě. Chromozom má uzavřenou strukturu, takže si ho zjednodušeně můžeme představit jako kružnici. Bakteriální chromozom je napojen na vnitřní stranu cytoplazmatické membrány většinou prostřednictvím mesozómu.
Stejně jako u jiných mikroorganismů má molekula DNA u bakterií tvar dvojité šroubovice. Ta je tvořená dvěma paralelními řetězci a ty jsou vzájemně spojeny vodíkovými můstky. Molekula DNA má jako základní stavební jednotku nukleotid. Ten se skládá z cukru deoxyribózy, z kyseliny fosforečné a dusíkaté báze. Dusíkaté báze máme purinové (adenin a guanin) a pyrimidinové (cytozin a tymin). Propojením deoxyribózy přes kyselinu fosforečnou s vnitřní bází přes vodíkové můstky principem párování (cytozin-guanin a adenin-tymin) vzniká polynukleotid. Dusíkaté báze mají přesné pořadí a určují složení bílkovin a ribonukleových kyselin v buňce. Vždy určitý úsek DNA představuje informaci o složení určité bílkoviny nebo ribonukleové kyseliny, anebo má určitou regulační funkci. Označujeme ho gen.
V klidové buňce bakterie je vždy jeden chromozom. Při rozmnožování dochází k zdvojení molekuly DNA neboli k replikaci, takže v buňce jsou dva chromozomy. Pak se buňka rozdělí ve dvě.
Poznámka
Řada bakterií obsahuje v buňce kromě chromozomální DNA ještě několik samotných molekul DNA uzavřené struktury, ale mnohem nižší molekulové hmotnosti. Nazývají se plazmidy. Některé z nich se uplatňují při spájení buněk (konjugaci) a mohou přenášet schopnost rezistence k různým antibiotikům nebo jiným jedům.
4.1.2.6
Bičíky
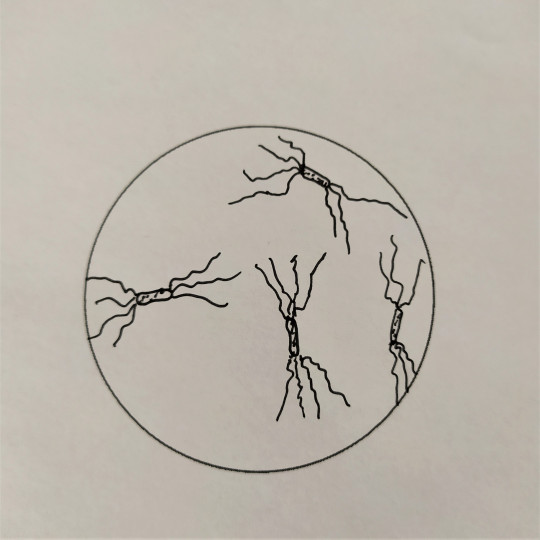
Některé tyčinkové bakterie mají schopnost se samostatně pohybovat, a to jim umožňují bakteriální bičíky. Bakteriální bičíky patří k povrchové struktuře bakterií. Jsou to dlouhá tenká vlákna, složená z bílkoviny flagelinu. Na dolní části mají háček a dvojici kotoučů, jimiž jsou zakotveny do cytoplazmatické membrány. Jsou většinou značně delší než bakteriální buňka. Podle počtu a umístění bičíků rozdělujeme bakterie na monotricha, lofotricha, amfitricha a peritricha. Bakterie, které nemají bičík, se nazývají atricha. Bičíky umožňují aktivní pohyb buňky ke zdroji živin, kyslíku apod. Hnací silou rotačního pohybu bičíků je elektrochemický potenciál vznikající na cytoplazmatické membráně v důsledku metabolismu buňky. Chemotaxe je pohyb bakterií do prostředí s optimální koncentrací určitých látek. Podobná reakce aerobních nebo anaerobních bakterií na kyslík se nazývá aerotaxe.
Poznámka
Na povrchu některých bakterií jsou umístěna dutá vlákna, označovaná jako pili nebo fimbrie. Vyskytují se u gramnegativních bakterií, a protože jsou adhezivní, vedou ke spojování buněk.
- atricha – bez bičíku
- monotricha – jeden bičík na jednom pólu těla buňky
- lofotricha – trs bičíků na jednom konci těla buňky
- amfitricha – trs bičíků na obou polárních koncích těla buňky
- peritricha – s bičíky po celém povrchu těla buňky
+
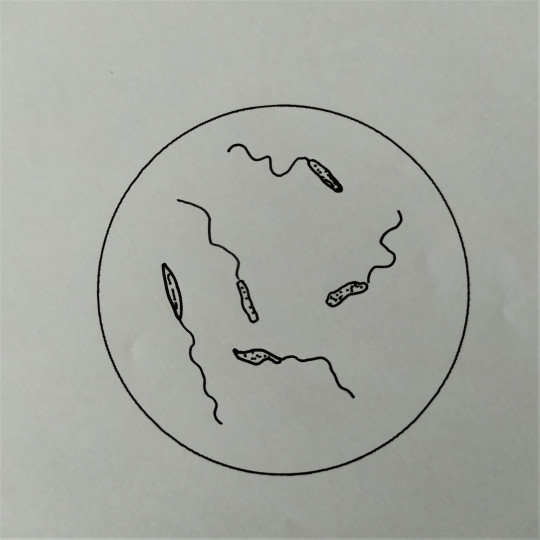
Obr. 25. Monotricha
+
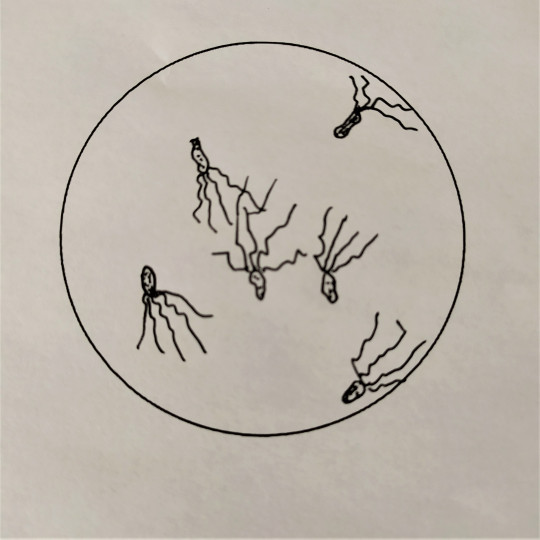
Obr. 26. Lofotricha
+
Obr. 27. Amfitricha
+
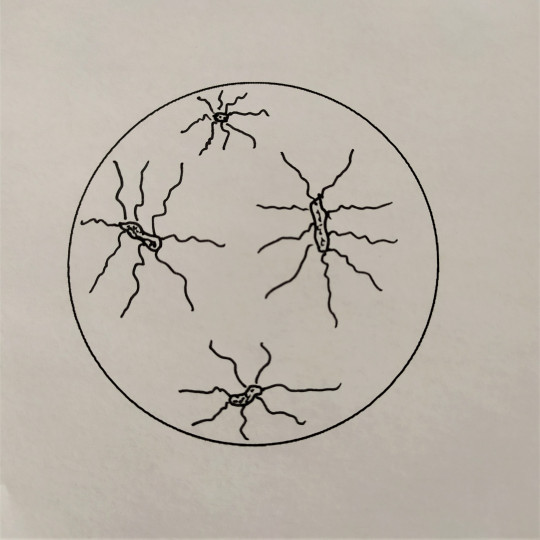
Obr. 28. Peritricha
4.1.3
Rozmnožování bakterií
Většina bakterií se rozmnožuje nepohlavně, příčným dělením. Příčné dělení probíhá tak, že mateřská buňka se prodlužuje ve směru podélné osy buňky až na dvojnásobek původní délky a její tloušťka zůstává stejná. Základem je tvorba přepážky, která vzniká z cytoplazmatické membrány a vede k rozdělení buňky na dvě části. Před rozdělením buňky dochází k replikaci chromozomální DNA. Jakmile má buňka určitou velikost a poměr buněčných bílkovin k DNA je na určité hodnotě začne probíhat replikace. Při rozmnožování bakterií se rozlišuje několik fází:
- G1-fáze, při níž je v buňce jen jeden genom a kdy probíhá syntéza bílkovin a ostatních buněčných složek,
- S-fáze, kdy se syntetizuje DNA a část genomu se vyskytuje již dvakrát, přičemž syntéza ostatních složek pokračuje,
- G2-fáze, při níž jsou v buňce dva genomy a začíná se syntetizovat přepážka oddělující mateřskou a dceřinou buňku.
Po ukončení replikace celé DNA se obě molekuly DNA oddělí a dochází k syntéze cytoplazmatické membrány, neboť chromozom je svým iniciačním bodem na ní napojen. Replikace DNA začíná v určitém iniciačním bodě a probíhá oběma směry podél celého chromozomu. Celková doba od vzniku dceřiné buňky k jejímu dalšímu rozdělení se nazývá generační doba. Je to doba, za níž dojde k zdvojnásobení počtu buněk a většinou také ke zdvojnásobení buněčné hmoty.
4.1.3.1
Konjugace u gramnegativních bakterií
Spájení buněk gramnegativních bakterií začíná spojením dvou buněk pomocí kanálku, který vytváří fimbrie. Kanálkem se přesouvá jedno vlákno chromozomální DNA z jedné buňky (tzv. donoru) do buňky recipietní. Vlákno, které v buňce chybí, se syntetizuje párováním dusíkatých bází. Kromě chromozomální DNA přechází do recipientu při konjugaci také plazmidy. Tak může recipient získávat nové vlastnosti. Spájení buněk jsou schopny pouze buňky bez plazmidů.
4.1.3.2
Spory bakterií
Některé grampozitivní bakterie mají schopnost tvořit uvnitř buňky buď kulovité, nebo elipsoidní útvary, které nazýváme spory. Spory jsou zvláštní útvary, velmi odolné proti nepříznivým vlivům vnějšího prostředí. Především zástupci rodů Bacillus a Clostridium. Vlastnosti spor jsou obdivuhodné. Jsou velmi odolné proti vysokým teplotám, dezinfekčním látkám, kyselinám a zásadám. Dokonce jsou odolné proti vysychání. Mají velmi silné a vodu nepatrně propouštějící povrchové pouzdro, které chrání uvnitř uložené pouzdro. Spora je umístěna buď uprostřed (centrálně) nebo na konci (terminálně). Spory některých druhů mají větší šířku než vegetativní buňka (jsou zduřelé). Podle tvaru a umístění rozpoznáváme čtyři typy spor (antraxový, subtilizový, klostridiový, plektridiový).
+
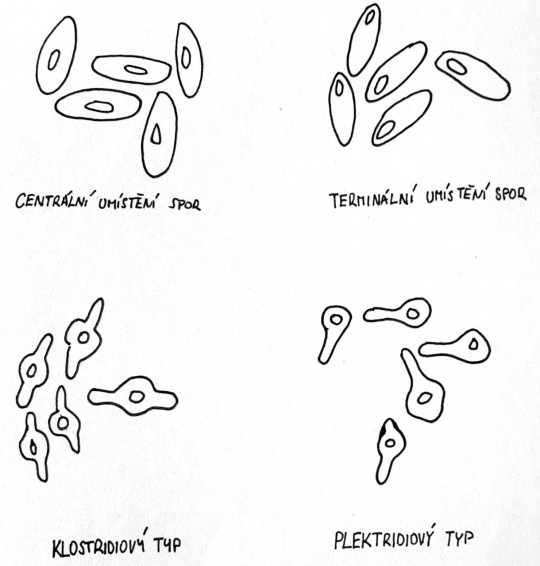
Obr. 29. Umístění spor
Vznik spor
Spora vzniká v situaci, kdy je dělení bakterie zastaveno většinou pro nedostatek živin. Buňka se protahuje a obsahuje dva chromozomy. Následují další fáze vzniku spory:
- Změna jaderného materiálu – oba chromozomy vytvoří jediné vlákno umístěné podélně.
- Vytvoření sporového septa – z cytoplazmatické membrány vzniká výběžek, který oddělí obsah cytoplazmy z mateřské buňky od budoucí spory a to včetně chromozomu, ribozomů, zásobních látek, enzymů a ostatních složek cytoplazmy.
- Vznik prespory – kolem oddělené části vznikají další dva obaly, vnější a vnitřní membrána.
- Tvorba obalových vrstev spory – mezi dvěma membránami vzniká kortex, ten se podobá peptidoglykanové vrstvě buněčné stěny. Mezi kortexem a vnitřní membránou vzniká další obalová vrstva, sporová stěna. Na povrchu se tvoří vnější plášť složený ze dvou vrstev bílkovinné povahy. U některých druhů se na povrchu vyskytuje povrchová blána (exosporium).
- Zrání spory – dochází k změnám uvnitř spory, ztrácí se voda až na 15 % obsahu, a vzniká dipikolinát vápenatý. Tyto změny jsou příčinou vysoké termorezistence.
- Uvolnění spory z buňky – vegetativní část bakterie se rozpadá a spora zůstává v prostředí.
Vlastnosti spor
Nejdůležitější vlastností spor je jejich termorezistence. Spory snášejí i několikahodinový var a ve vlhkém prostředí jsou usmrcovány až při 115 - 120 °C po dobu 15 - 30min. V kyselém prostředí se tato odolnost snižuje. Příčinou termorezistence je nízký obsah vody, nahromadění dipikolinátu vápenatého, kortex a vysoký obsah lipidů v obalových vrstvách. Další důležitou vlastností je zvýšená odolnost k jedovatým látkám a to díky nepropustnosti obalových vrstev. Spory pro zvýšený obsah sirných aminokyselin v bílkovinných obalech a pro absenci volné vody mají i mírně zvýšenou radiorezistenci. Další vlastností je zvýšená rezistence k vysychání, hladovění, a jiným nepříznivým podmínkám z důvodu nízké metabolické aktivity. Spora je klidovým stavem bakterie.
Klíčení spor
Přeměna spory ve vegetativní buňku nastává po přenesení spory do vhodných podmínek. Musí mít dostatek živin, vody, vhodné pH a teplotu. Postupně se rozpadne kortex a uvolní se organické sloučeniny do prostředí. Zároveň začne spora přijímat vodu a dojde k prasknutí obalů. Spora se stane vegetativní buňkou.
+
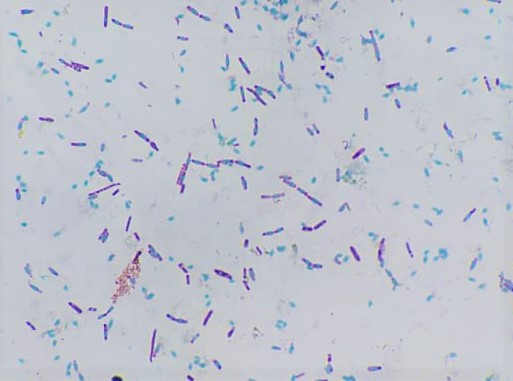
Obr. 30. Mikroskopický preparát – spory Bacillus cereus